
BY ELLIOT SMITH
Winning Entry for The Huxley Prize 2025
Ask almost any biology student around the country what the mitochondria are and it’s likely they will recite the mantra: they’re the powerhouse of the cell. It’s a phrase so overused it has become a punchline, yet it holds much truth. The organelles harness the power of the chemiosmotic gradients it creates, channelling the flow of protons through the protein complex ATP synthase, driving ATP synthesis in a process that generates a potential difference across the membrane in the order of ‘150 to 200’ millivolts. The membrane is so thin (around 6nm) that the intensity of the electrical field you would experience within the membrane region is around ‘30 million volts per metre, equal to a bolt of lightning’. Without this energy supply, complex life would grind to a halt. However, this is only half of the story, for these same organelles, so key in sustaining life, also hold the power to end it.
Apoptosis (from the Greek for ‘falling off) is a very precise and carefully choreographed mechanism for cellular suicide. Although this sounds somewhat malicious, the process is crucial for growth, health and survival in our bodies, and is a principle that runs through life in all its forms. Billions of our cells succumb to this fate every day for the greater good, eliminating defective or dangerous cells, maintaining tissue homeostasis and preventing potential cancers from forming. But when the regulation of apoptosis falters, the consequences can be devastating. Too little cell death can lead to uncontrolled growth (cancer), while too much can cause degenerative or autoimmune diseases through the wiping out of perfectly healthy cells.
Yet the way in which apoptosis is carried out raises an evolutionary mystery. The conventional view frames apoptosis as an altruistic act of cellular suicide – a mechanism devised by the organism that cells use to contribute to maintaining the health of the larger organism. In this interpretation, over evolutionary time, our cells have evolved self-destruct sequence to remove themselves, when damaged or unneeded. But an alternate hypothesis casts the mitochondria in a more sinister light. In his book ‘Power, Sex, Suicide,’ author Nick Lane argues that mitochondria’s central role in apoptosis may trace back to their bacterial origins billions of year ago, before they became fully integrated into the host cell. If this view is correct, this may suggest that apoptosis is an inherited murder weapon, taken from our cellular invaders.
By pointing out that apoptosis often starts with the release of mitochondrial inner membrane proteins (such as cytochrome C) and the fact that many key apoptosis-inducing factors have clear bacterial roots, Lane alludes to the possibility that this lethal machinery may once have been used by the mitochondria to selfishly murder its host cell, rather than to serve it.
Understanding where apoptosis came from is not just a historical curiosity. Its bacterial origins can explain why the mitochondria, rather than the nucleus, function as the judges of cellular fate, whilst also gaining crucial insight into both the evolution of complex life and the mechanisms modern medicine could exploit to shift the balance of life and death in diseased cells.
In the sections that follow, I will first explore the molecular machinery of apoptosis, investigate the early symbionts that lead to origin of the eukaryotic cell and complexity, and consider the ‘suicide vs murder’ evidence, attempting to answer the essay’s central question.
Early Discovery
For more than a century beforehand, scientists had been familiar with necrosis – the chaotic, unregulated death of cells caused an infection, injury, or other damage. When looking down a microscope, ‘necrotic’ cells swell, rupture and spill their contents, causing inflammation in the surrounding tissue.
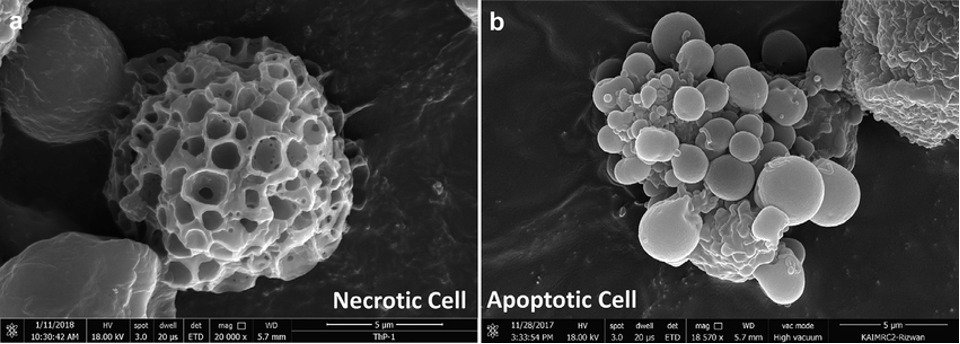
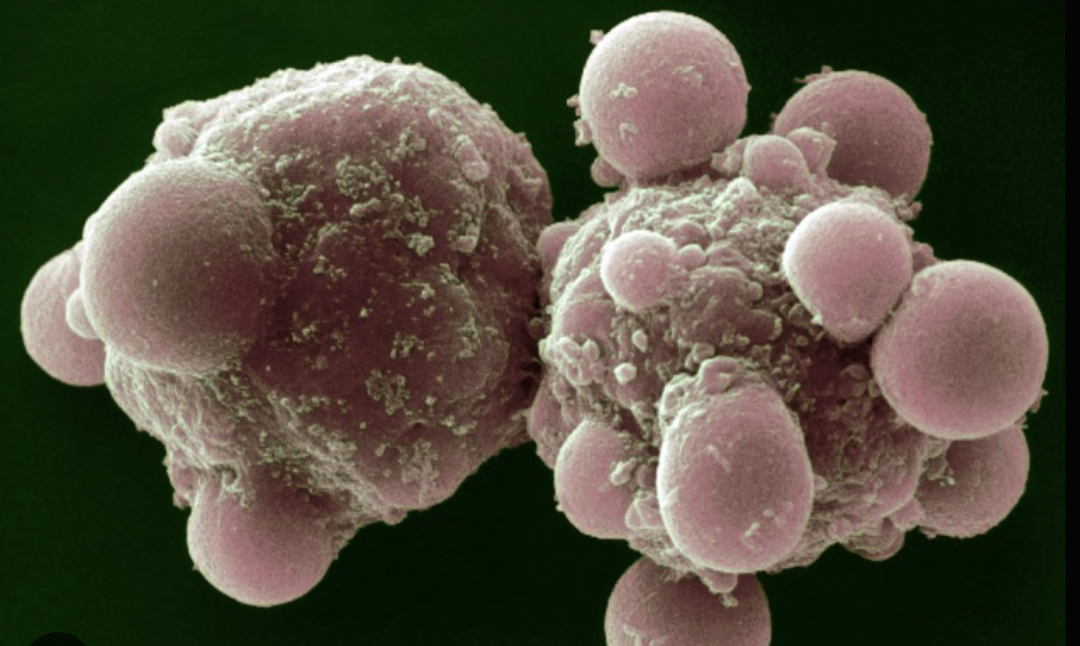
However, in 1972, a landmark paper written by John Kerr, Andrew Wylie and Alastair Curie coined the term ‘apoptosis’ – a controlled form of cell death. In contrast to necrosis, apoptotic cells shrink, condense their chromatin and break into membrane-bound ‘apoptotic bodies’ (seen on the right in the image above) without causing any inflammation.
Wyllie and his colleagues therefore recognized apoptosis as a regulated ‘suicide’ mechanism, crucial for tissue homeostasis. This work established clearly that cell death wasn’t just a passive collapse, but could be an active, gene-directed process. At the time, this concept was revolutionary and thus set the stage for apoptosis to be studied for many years to come.
However, the major turning point came in the mid-1980s when H. Robert Horvits and his colleagues at MIT set about identifying the genes responsible for apoptosis in a tiny nematode worm. Caenorhabditis elegans, microscopic in scale, was in many ways the perfect candidate for the research. Firstly, it was transparent, allowing for researchers to track the fate of individual cells. Secondly and more importantly, a predictable group of cells, 131 out of the 1090 body cells that comprise the nematode, die by apoptosis during embryonic development with extraordinary invariance.
Through further research, Horvitz identified the first ‘death genes’ by discovering that mutations in these genes (ced-3 and ced-4) would prevent the 131 cells from dying. Put simply, the genes were required to function properly in order to execute the cell-death programme. But the more fascinating and unexpected discovery was that there were exact equivalents of these genes in mammals, plants and even flies, so closely related that they could be genetically engineered to replace the nematode genes in the worms with no issues.
This discovery had huge implications in areas such as cancer research: if these same mutations occurred in humans, then the growing cancer cells would likewise to fail to commit suicide and instead proliferate, forming into a tumour. This groundbreaking work earned Horvitz and his team the Nobel Prize in 2002, whilst also establishing apoptosis as a universal biological mechanism.