
BY ELLIOT SMITH
Winning Entry for the Huxley Prize 2025
The Molecular Machinery of Apoptosis
The central players in apoptosis are a specialised family of proteases known as the caspases – the cell’s executioners. Discovered as the functional equivalents of the worm’s ced-3 gene, the caspases are continually produced in the body, waiting in an inactive state, poised and ready to jump into action. Once activated, the caspases dismantle the cell with extreme efficiency, slicing up structural proteins supporting the cytoskeleton, enzymes involved with DNA repair, degrading the cell and its components from the inside.
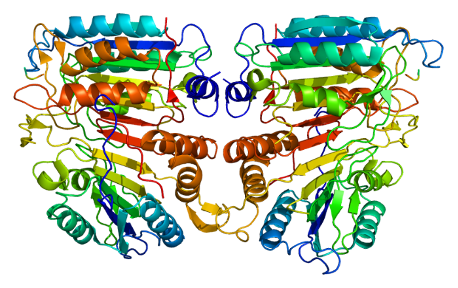
The caspases fall into two broad classes: the initiator caspases and the executioner caspases make up the eleven known members in humans. The initiator caspases, such as caspase-9 (seen on the right) and caspase-8, do exactly what you would expect from their title: initiating the death programme. They function as sensors, responding to the arrival of a death signal, triggering the executioner caspases and setting the process in motion. The executioner caspases (most notably caspase-3 and caspase-7) are the cell’s demolition crew. This division of labour means that once a very small number of initiators are activated, they can switch on and activate hundreds of executioners, which, momentarily after, cleave thousands of proteins throughout the cell. This amplification transforms the small signal to an unstoppable force leading to the cell’s eventual death.
All of this results in the structured choreography Andrew Wylie first described under his microscope: the cytoplasm contracting as actin filaments are severed, chromatin condensing and its DNA cleaved into regular fragments and the cell surface membrane bulging before ‘blebbing’ into small membrane-bound apoptotic bodies. These apoptotic bodies consist of cytoplasm containing tightly packed organelles with or with or without fragments of the nucleus. By no means are these bodies wasted, forming neat parcels of the cell’s contents, engulfed by neighbouring cells or macrophages, therefore avoiding the spillage of inflammatory signals into the surrounding tissue.
The alternative to apoptotic cell death is necrosis. In contrast to apoptosis, necrosis is a chaotic, passive collapse that arises from overwhelming stress or damage. As ATP levels fall, the cell loses its ability to control and regulate its internal environment. Ions and water rush in, the cell swells, organelles rupture and the plasma membrane eventually breaks open, provoking an inflammatory response. While this response is meant to prevent further consequences, it often harms healthy neighbouring cells, causing pain, swelling and in some cases permanent damage.
Mitochondria: Powerhouse to Executioner
For many decades, biologists around the world searching for the control centre of apoptosis assumed the orders came from the nucleus. Since it houses the genome and the life/death genes we talked about earlier- this was a perfectly reasonable guess. Yet in the mid-1990s, a series of surprising discoveries revealed the key orchestrator was not the nucleus at all, but the mitochondria.
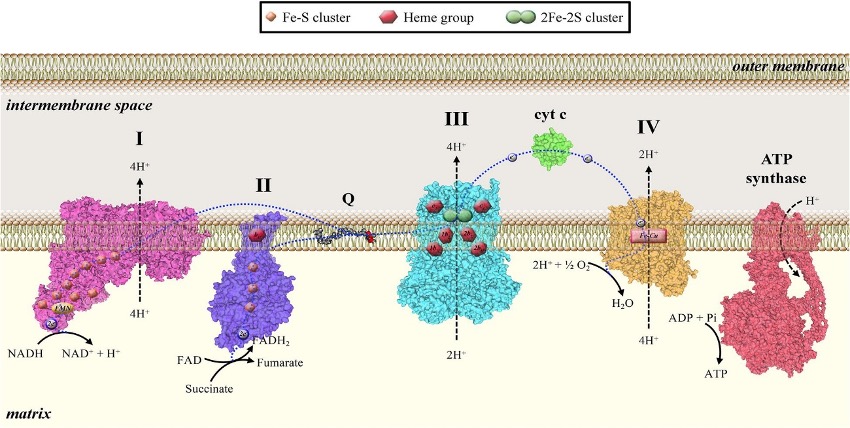
This realization dawned when researchers found that a typically harmless mitochondrial resident: cytochrome c is released into the cytoplasm to trigger apoptosis. Under normal conditions, cytochrome c – a single-chain globular protein – acts an electron carrier in the electron transport chain. More specifically, it facilitates the transfer of electrons from complex three to complex four (see above), aiding in the generation of the proton gradient across the inner mitochondrial membrane that powers life. The indispensability of cytochrome c becomes clear during certain studies: the selective elimination of cytochrome c in adult forebrain neurons leads to severe brain abnormalities and premature death, highlighting its vital role in maintaining tissue homeostasis. But this same molecule, when displaced from its home, switches roles entirely.
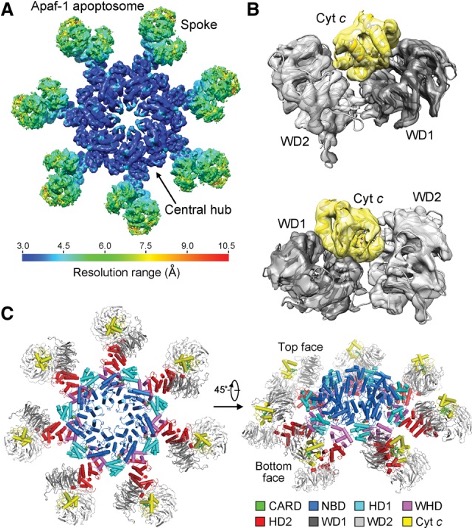
How does the release of this small, seemingly benign molecule, lead to the death of a cell? When a cell receives a distress signal, from irreparable DNA damage for instance, mitochondria respond by becoming permeable, a step know as mitochondrial outer membrane permeabilization (MOMP for short). Cytochrome c is normally tethered to the outside of the inner mitochondrial membrane, but due to this permeabilization, the membrane develops pores, allowing cytochrome c and other pro-death factors to flood into the cell’s cytosol. Here, cytochrome c binds to a protein called Apaf-1, assembling a mega-complex named the apoptosome.
The apoptosome is effectively a molecular death star, and recruits and activates caspase-9, which in turn triggers proteases like caspase-3, bringing about the controlled collapse of the cell. The figure shows the human apoptosome – a wheel-shaped ring, in all its geometric glory, with a central hub and seven neat spokes. When cytochrome c docks into the ‘spokes’, the ring snaps into its functional state, bringing caspase-9 (for example) molecules together into the hub activating each other, before the signal passes onto caspase-3 to carry out the orderly shutdown.
It’s worth noting that mitochondria unleash a whole arsenal for this task, not just cytochrome c. Factors like Smac/DIABLO can be used (which neutralize the inhibitors for death), as well as AIF (apoptosis inducing factor) which can trigger a form of death even when the caspases are restricted. In summary, the mitochondrion, serves as the decision maker and trigger for the cell’s apoptotic programme.