
BY ELLIOT SMITH
Winning Entry for the Huxley Prize 2025
The Bcl-2 Family
Mitochondria clearly hold the power to end a cell’s life with superb efficiency, yet cells don’t just self-destruct at the first hint of discomfort. Everyday stresses such as a short burst of a reactive oxygen species or a temporary nutrient dip are routine parts of cell life. So how does a cell know when to endure and carry on, and when to accept its fate of death? What prevents every flicker of stress from initiating the apoptotic response?
The answer is another group of proteins known as the Bcl-2 family. These proteins are split into two groups: those that promote the cell’s survival and those that press for its destruction. In periods of health, the protectors – proteins like Bcl-2 and Bcl-xL – keep the mitochondrial pores sealed, preventing the exit of molecules such as cytochrome c. But when the balance tips the other way and cellular health declines, the enforcers start to take charge. Proteins Bax and Bak can punch holes into the mitochondrial membrane, allowing the death factors to escape.
As this happens, the membrane potential collapses. The extraordinary voltage that mitochondria normally sustain across their barriers, a few nanometres thick, suddenly dissipates. Alongside this, the electron transport chain falters, and electrons leak to oxygen leading to a burst of oxygen free-radical generation. These reactive oxygen species not only damage nearby DNA and proteins but also amplify the death cascade.
Therefore, the balance between these two opposing factors of the Bcl-2 family is what determines the fate of the cell. If this balance tips towards the pro-apoptotic side, the membrane becomes permeable and apoptosis proceeds. The Bcl-2 proteins form a molecular council of sorts, voting on life or death of the cell. This council listens to signals from every corner of the cell. DNA damage, loss of growth factors, bursts of oxidative stress can all sway the vote. If enough of these warnings pile up, the mitochondrial gates open and the cell is committed to self destruction.
It is worth quickly noting that apoptosis is not only triggered from within. There is also a second route- the extrinsic pathway- where orders come from the outside. For example, death signals such as ‘Fas ligand (FasL)’ bind to receptors on the cell surface and switch on the initiator caspases directly. Additionally, some death signals alter the balance of the feuding protein family in favour of apoptosis. Thus, the bcl-2 proteins integrate a diverse range of signals from both inside and out of the cell, calibrating their strength in the mitochondria.
Now that we know how apoptosis works at the molecular level, it is time to face the question at hand. Was this programme born out of cooperation, or did it begin as a deadly strategy in the conflict between an archaeal host and its newly arrived α-proteobacterial residents (later mitochondria)? On one side of the argument, the hydrogen hypothesis and other models explaining the early symbiosis suggest the partnership was mutually beneficial from the outset, driven by metabolic exchange. On the other side, the striking similarity between today’s apoptotic molecular machinery and bacterial toxins begs the question: did the early mitochondria kill their host cells in their own interests? If this view is correct, then apoptosis is better seen as murder than as suicide.
This view may sound unusual and unfamiliar at first, but some argue the roots of apoptosis lie not in mutual benefit, but in an ancient struggle for survival. The best way to approach this paradox is to rewind to the very start, around two billion years ago, at the dawn of eukaryotic life and its first symbionts.
Early Symbiosis: The Hydrogen Hypothesis
For almost two billion years, life on earth was small, simple and single-celled. The prokaryotes (single-celled organisms lacking a nucleus or internal compartments): bacteria and archaea, which first appeared at least 3.5 billion years ago, reigned supreme across almost every environment the planet could offer. Archaea, though they look similar to bacteria under a microscope, are a distinct domain of life with unique cell membranes, genetic machinery and metabolic pathways that allow them to thrive in the most extreme of environments.
However, for all their versatility, conquering oxygen-free muds, salty lagoons and boiling vents, no bacterium or archaeon ever made the leap to true complexity- they developed no nuclei, no complex internal compartments and, most crucially, no multicellularity. Then, around two billion years ago, an extraordinary event occurred: an archaeal host cell formed an alliance with a bacterium, and it was from this that the first eukaryotic cell was born.
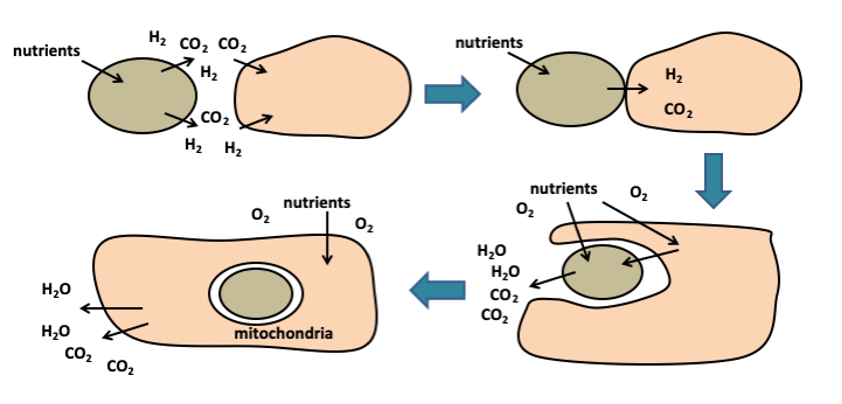
The best explanation for how this alliance formed is through the hydrogen hypothesis, proposed in 1998 by William Martin and Miklos Muller. In this model, the host cell was a strictly anaerobic archaeon – most likely a methanogen- which lived by combining hydrogen with carbon dioxide to make methane. This process, known as methanogenesis, proceeds via the reaction:
4H₂ + CO₂ → CH₄ + 2H₂O
The reaction releases enough energy for the archaeon to synthesis ATP and survive in oxygen free environments. Therefore, its survival depended entirely on hydrogen.
The problem came when hydrogen in the environment became harder to come by. Around 2-1.5 billion years ago, as oxygen began to accumulate in the oceans and atmosphere, sulphate became more abundant, fuelling the rise of sulphate-reducing bacteria. These microbes were ruthless competitors, consuming the hydrogen before the methanogens could get to it, presenting an energy crisis to the methanogen.
The solution arrived in the form of a bacterium: an α-proteobacterium that conveniently produced hydrogen, a waste product from its metabolism. For our archaeal host, this was like liquid gold and, by cosying up to the bacterium, the host secured a private hydrogen supply that it could use to fuel its energy synthesis. For the bacterium, life inside the host meant a stable environment, rich in nutrients where it could prosper.
At first this may have been a loose, external partnership with the host and symbiont nestled side by side in oxygen-poor sediments. However, over time, the bacterium were engulfed into the methanogen, transforming the relationship into an endosymbiotic one, in which one organism lived inside the other in a mutually beneficial partnership. Once internalised, the α-proteobacterium became the common ancestor of all mitochondria. In this way, unlike older theories that imagined the mitochondria being engulfed long after the establishment of the eukaryotic cell, the hydrogen hypothesis insists that the act of bringing in the bacterium created the eukaryotic cell. Besides, it’s not a coincidence that all known eukaryotes either have mitochondria or show evidence of having them at one point. From this one improbable encounter emerged the Last Eukaryotic Common Ancestor (LECA), the origin of all complex life.
The payoff was revolutionary: by internalising a bacterium capable of respiration, the archaeal host gained access to an energy supply vastly greater than any free-living prokaryote could manage. Prokaryotes are limited by their surface membranes; there is only so much ATP they can make. This is because ATP is always synthesised across a membrane, where proton gradients drive the turbines of ATP synthase, and prokaryotes have only their outer cell membrane to work with. With the addition of the primal mitochondria, the archaeal cell could spread its respiration across thousands of internal membranes, scaling up energy production immensely.
This surplus energy drove a series of innovations that were energetically out of reach for prokaryotes: the maintenance of far larger genomes packaged inside a nucleus, the evolution of a dynamic cytoskeleton and the energy costs of sexual reproduction. As Martin, Muller and later Lane emphasise, mitochondria did not merely power life as we know it – they made it possible.
Yet, alongside this gift of energy, the new lodger also carried with it a darker legacy. The same membranes and molecular tools that enabled respiration could, under contrasting circumstances, could be turned against the host methanogen. Indeed, many of the key factor that drive apoptosis today bear an uncanny resemblance to bacterial toxins. This raises the provocative question: was the endosymbiont only a benevolent partner, or did it once, long ago, harbour the tools to sabotage its host from within.