
BY ELLIOT SMITH
Winning Entry for the Huxley Prize 2025
The Parasitic Origins of Programmed Cell Death
It may seem slightly counterintuitive that the very process cells use for the greater good of the organism may have begun as an act of selfish sabotage. Yet some argue exactly this: apoptosis might trace back to proto-mitochondria behaving as internal parasites. In his book, Lane raises the possibility that the opportunistic invaders ‘killed their host cells in their own interests,’ making apoptosis ‘better seen as murder than suicide.’
Modern evidence does in fact strongly support this once radical idea. Through comparing DNA sequences and protein structures across species, scientists have been able to trace their evolutionary origins. This analysis has shown that all of the key components of our apoptotic machinery can be matched back to the genes and proteins of bacteria, rather than to the archaeal host.
For instance, cytochrome c which we met earlier – the trigger which unleashes the caspase enzymes, is encoded by a gene whose closest relatives are found in α-proteobacteria, the bacterial group from which mitochondria evolved, rather than the host cell. In fact, with the only exception being AIF (apoptosis-inducing factor), all known apoptotic proteins released from the mitochondria are bacterial in origin and are absent from the archaea. This includes notorious death factors like ‘Endonuclease G’ and the DNA slicing caspases themselves, whose closest cousins are found in bacteria. Even the cell’s pore-forming Bcl-2 family bear a striking resemblance to bacterial toxins. Their 3D structure mirrors that of certain bacterial poisons (like the diphtheria toxin) that drills holes into host cell membranes.
Kaushal et al. (2023) reinforced this view in a study showing that when yeast (eukaryote) apoptotic genes were replaced with equivalent genes and proteins from bacteria (prokaryote), the foreign versions still triggered cell death under stress. This interchangeability implies the lethal function of these proteins have been conserved since the early symbiosis, making it far more like that apoptosis was inherited from early bacteria. The alternative – that the same complex machinery evolved twice, once in bacteria and again in eukaryotes – would require an improbable case of convergent evolution and is far less probable.
It appears, therefore, that the proto-bacteria brought with them the arsenal of such proteins, which could rupture membranes and disrupt host cells from within, supporting the notion of ‘murder from within, rather than suicide.’
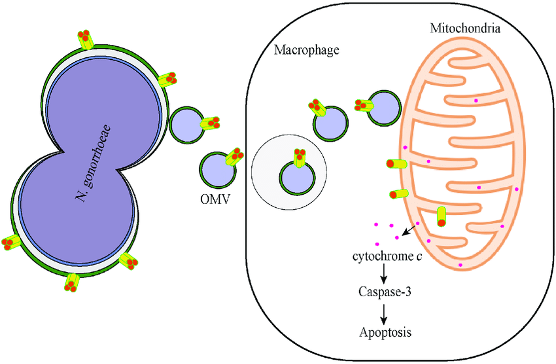
In 1997, José M. Frade and Theologos Michaelidis presented this concept in its first clear form. They pointed to modern bacteria like Neisseria gonorrhoeae – the parasite causing the STD gonorrhoeae with a devious host-killing strategy, as a living model of how the early mitochondria may have behaved. N. gonorrhoeae inserts a protein pore (PorB) into the membranes of its host cell. As long as the host cell stays metabolically healthy, the pore remains plugged by the ATP molecules that the host cell synthesises. But, if the host’s energy levels falter, the PorB channels open, causing a detrimental depolarisation of the mitochondrial membrane and triggering the cell’s apoptosis programme. In this way, the bacterium provokes the cell to destroy itself and subsequently lives off the remains of the host, escaping to infect new cells.
Michaelidis argued that the proto-mitochondria could have deployed a similar survival mechanism: lying in wait within the host methanogen, monitoring its health, and inducing host cell suicide whenever the host became compromised. Notably, some other invasive bacteria today (such as some Rickettsia species) follow this exact pattern, lending to the idea that the first mitochondria were domesticated killers.
It might seem strange that early cells tolerated such a potentially dangerous lodger at all. From an evolutionary logic standpoint, it does in fact make sense. For the invading bacterium, having a kill-switch of sorts was a sensible strategy: if the host cell became unhealthy or starved, it could pull the plug and escape to a new host, preventing the parasite going down with the host. For the host, this was at first disastrous – until it found a way to turn the tables. Over time, the ancestral host cell may have evolved its own regulators to control and contain its parasite. Proteins like the Bcl-2 family may have arisen to bind the pores and prevent the release of the death factors, taming the would-be killer. In effect, the host cell domesticated the lethal apparatus, eventually recruiting it for its own cellular quality control: a crucial policing tool once cells began living in multicellular communities.
This argument leaves us with a striking paradox. When we looked earlier at the origin of eukaryotes, we framed it as a peaceful metabolic partnership rather than a parasite war. Nothing in this field is certain – these events happened two billion years ago, leaving no direct record. All we have are molecular echoes and present-day counterparts, leaving it up to us on how we weigh the fragments of evidence. The challenge therefore is to decide which picture carries more weight and whether the evidence for a parasitic origin of apoptosis really outweighs the case for a peaceful symbiosis. To do this, we must now turn to the evidence against the ‘murder’ argument.
Counterarguments to the ‘Murder Weapon’ hypothesis
The proposition that apoptosis began as a bacterial weapon is provocative, but the case is far from settled. It is clear that mitochondria supplied most of the death machinery and were instrumental in apoptosis’ evolution. And yes, some of the proteins that control apoptosis in eukaryotes, like the Bcl-2 family that regulate the mitochondrial pores, do resemble pore-forming toxins in some modern-day bacteria. Yet, this similarity may have arisen simply as a result of convergent evolution: when faced with similar challenges of puncturing membranes, natural selection tends to produce similar shapes and mechanisms, because those designs work best. Although there are clear parallels between our cell death machinery and bacterial toxins, this may not prove inheritance or evolutionary relationship.
Another difficulty lies in explaining how a parasite could deliver a complete cell-death program into its host without destroying itself in the process. The modern pathogens (we mentioned these earlier) can trigger apoptosis in human cells by inserting their own porins, but rely on the host’s existing machinery to conduct the process itself; they are flipping a switch and do not carry the whole functional system. For proto-mitochondria to enforce death from within, they would have needed a self-contained murder kit and a way to deploy it without self-destruction, since any pore-forming toxins could just as easily have ruptured their own membranes. Although this may have been possible, many are sceptical that such a complex package could have been present in the earliest of endosymbionts, particularly given that no modern bacterial relatives are known to possess a complete set of machinery.
A further consideration for the murder weapon hypothesis is evolutionary backdrop. Apoptosis is crucial in multicellular organisms, where the wellbeing of the entire body depends on the regulation of the behaviour of individual cells. But these particular pressures barely existed for single-celled ancestors, which did not need to sacrifice themselves as the larger organism didn’t exist. If apoptosis was passed down in its entirety from the proto-mitochondria, we would expect to see the same machinery across all eukaryotes; however, this is not the case. Some display features that resemble apoptosis, such as DNA fragmentation, but they achieve this using different proteins rather than the caspases seen in plants. Plants, for example, use metacaspases – enzymes that are unrelated in their ancestry to caspases but adapted to fulfil a comparable role. The fact that such distinct pathways have produced the same outcome suggest that programmed cell death has been pieced together independently in different lineages.
The murder weapon argument also sits uneasily alongside the hydrogen hypothesis, the theory we mentioned earlier which explains the origin of eukaryotes as a metabolic partnership. The hydrogen hypothesis emphasises mutual benefit from the outset: the archaeon consuming the hydrogen produced by the bacterium. By contrast, the idea of mitochondria arriving as parasites armed with toxins suggests a relationship consisting of conflict rather than of cooperation. A parasite can gain by killing its host and escaping to a new one, but in the case of a symbiont reliant on its partner’s chemistry, there is not much to gain by destroying its source, especially if it had no means of finding another one. Although some α-proteobacteria are motile, the chances of it successfully leaving and finding another suitable host would have been extremely slim, especially as it became increasingly more genetically dependent on its host.
One plausible scenario is that the mitochondria provided basic ingredients – molecules like cytochrome c and pore-forming factors: signals or triggers of death. These could have been integrated by the host cells into their own regulatory systems shaped by evolutionary pressures. The range of these molecular pathways may point to repeated reinvention, with each one using mitochondrial components in different ways to create similar cell death machinery.
Conclusion
Apoptosis embodies one of biology’s biggest paradoxes: a fundamental process of programmed cell death that sustains life. At the smallest of scales, it looks like an altruistic act, yet the deeper we go into its origins, the more the tools used in apoptosis shows a clear bacterial ancestry.
In this essay we have examined apoptosis, from its discovery to the enzymes that dismantle the cell, to the bacterial fingerprints that hint at its ancient origins. We have weighed the two competing stories – one presenting it as altruism, the other as ancient attack. The truth, however, is elusive. The fact these events unfolded two billion years means the only traces of the answer left are the scattered molecular clues that need to be pieced together. We cannot simply rewind the tape of evolution to see whether early mitochondria really killed their hosts; the best we can do is weigh the hints that remain.
Origins aside, apoptosis is a fascinating example of how evolution has reshaped a force of destruction into one of control. The same processes that may once have ruptured the membranes of the host cell from within are now indispensable for the body – maintaining balance, shaping growth and policing cancer.
References:
Lane, N. (2018) Power, sex, suicide: mitochondria and the meaning of life. 2nd edn. Oxford: Oxford University Press.
Lane, N. (2015) The vital question: why is life the way it is? London: Profile Books.
Mustafa M, Ahmad R, Tantry IQ, Ahmad W, Siddiqui S, Alam M, Abbas K, Moinuddin, Hassan MI, Habib S, et al. Apoptosis: A Comprehensive Overview of Signaling Pathways, Morphological Changes, and Physiological Significance and Therapeutic Implications. Cells. 2024; 13(22):1838. https://doi.org/10.3390/cells13221838
Elmore, Susan. “Apoptosis: a review of programmed cell death.” Toxicologic pathology vol. 35,4 (2007): 495-516. doi:10.1080/01926230701320337 Apoptosis: A Review of Programmed Cell Death – PMC
Vandana Kaushal, Joanna Klim, Adrianna Skoneczna, Anna Kurlandzka, Tuguldur Enkhbaatar, Szymon Kaczanowski, Urszula Zielenkiewicz, Apoptotic Factors Are Evolutionarily Conserved Since Mitochondrial Domestication, Genome Biology and Evolution, Volume 15, Issue 10, October 2023, evad154, https://doi.org/10.1093/gbe/evad154
Martin, William & Muller, Miklos. (1998). Martin, W. F. and M??ller, M.. The hydrogen hypothesis for the first eukaryote. Nature, 392: 37-41, 10.1038/32096. Nature. 392. 37-41. 10.1038/32096. https://www.researchgate.net/publication/51306904_Martin_W_F_and_Mller_M_The_hydrogen_hypothesis_for_the_first_eukaryote_Nature_392_37-41_10103832096
Zhou, Zhuan et al. “Diverse functions of cytochrome c in cell death and disease.” Cell death and differentiation vol. 31,4 (2024): 387-404. doi:10.1038/s41418-024-01284-8 Diverse functions of cytochrome c in cell death and disease – PMC